Представлена версия документа, предназначенная для печати
Вы можете загрузить эту страницу на сайте
Под влиянием антропогенного фактора происходит уменьшение числа видов аборигенной фракции флоры и увеличение числа видов за счет роста адвентивной фракции. Рассмотрено изменение систематической структуры локальных флор с учетом адвентивной фракции вдоль широтного градиента. Участие адвентивных видов слабо меняет таксономический спектр флор. В целом антропогенное воздействие на флору приводит к обеднению разнообразия флоры, не меняя ее структуры и основных закономерностей распределения. |
Антропогенное влияние на разнообразие флоры имеет две основные тенденции: уменьшение числа видов и/или изменение видового состава аборигенной фракции флоры и увеличение числа видов за счет роста адвентивной фракции. Адвентивными являются виды, проникновение которых связано с деятельностью человека. Их расселение происходит либо путем случайного (непреднамеренного) заноса, либо в результате дичания культивируемых растений [12].
В ряде работ отмечено, что процент исчезнувших аборигенных видов обратно пропорционален размеру территории и прямо пропорционален степени действия антропогенного фактора. Так, при изучении локальных флор южной части Приволжской возвышенности (Саратовская область) доля исчезнувших видов оказалась выше для флоры небольшого поселка (8.6%), а не крупного города (5.1%) [28]. В первую очередь под влиянием антропогенных факторов исчезают: а) редкие виды и виды, находящиеся на границе своего ареала, б) водные виды, виды влажных местообитаний и сегетальные сорняки [3].
Факторы антропогенного воздействия на флору довольно многообразны, однако их влияние на характер и скорость изменения флоры изучено еще недостаточно. Сокращение числа видов прежде всего связано с уничтожением их местообитаний. Наиболее значительное влияние на сокращение жизненного пространства существования вида оказывает распашка территории [3, 6, 28]. Среди других факторов часто отмечается осушение болот и связанная с этим процессом добыча торфа, которые приводят к уничтожению многих видов болотной флоры [22]. Выпас скота и сенокошение прежде всего влияют на произрастание луговых видов. Исчезновение видов обусловлено также эвтрофикацией территории [3], урбанизацией ландшафтов и увеличением рекреационной нагрузки, а также сбором лекарственных и декоративных растений в окрестностях крупных городов [13, 22], строительством и техногенными разработками [9].
Вторая составляющая антропогенного воздействия связана с появлением новых для конкретного региона, адвентивных видов. Этому вопросу посвящено наибольшее количество работ, в которых исследуется антропогенная трансформация биоты. Изменение числа адвентивных видов объясняется как действием отдельных факторов, так и особенностями характера биоты. Для флоры в целом следует отметить следующие моменты.
Изучение распределения адвентивных видов осложняется еще и тем, что две составляющие антропогенного воздействия проявляются независимо друг от друга и зачастую обусловлены разными переменными. При синантропизации флоры четкая тенденция увеличения флористического богатства наблюдается только на первых этапах антропогенного воздействия на флору, на последующих этапах происходит заметное уменьшение общего числа видов во флорах, как за счет аборигенных видов, так и за счет адвентивных [3, 32].
В литературе неоднократно высказывалось предположение, что одним из следствий изменения флоры под действием антропогенных факторов является ее унификация, «внедрение адвентивных видов маскирует и обедняет специфичность флор» [17, с. 13]. Однако, большинство работ, посвященных этому вопросу [1, 18], основано на анализе состава и структуры синантропной флоры, т. е. флоры антропогенных местообитаний, которая формируется как из заносных, так и из местных видов, причем последние преобладают.
Вместе с тем крайне мало внимания уделяется анализу систематической структуры флор с учетом адвентивных видов. Подробные исследования проведены в небольшом числе регионов [10, 28]. На глобальном уровне было продемонстрировано, что распределение семейств адвентивной флоры соответствует регионам, занимаемым семействами естественной флоры [36]. Некоторыми авторами высказано предположение о том, что структура сформировавшейся адвентивной фракции частично соответствует структуре аборигенной фракции флоры той же территории [3].
Настоящая работа посвящена анализу структуры естественных флор с учетом адвентивных видов и имеет несколько целей: 1) проследить изменение числа адвентивных видов вдоль широтного градиента; 2) оценить изменения систематической структуры флор с учетом адвентивных видов; 3) провести анализ зависимости числа адвентивных видов от уровня видового богатства флор.
Изменение числа заносных видов вдоль широтного профиля изучено по данным 20 локальных флор, распределенных с севера на юг вдоль меридионального сектора Восточной Европы от южных тундр до зоны степей, а таксономическая структура – на основании 17 локальных флор (табл. 1, 2). Одна из проблем, возникающая при сравнении флор из различных регионов, связана с унификацией видовых списков. Поэтому изучение участия адвентивных видов проведено с использованием базы данных, представляющей местонахождения видов в конкретной точке и имеющей единый номенклатурный список видов, основанный на сводке С. К. Черепанова [30]. База данных (Alien plant Species) составлена с помощью СУБД Paradox 7.0 for Windows, и состоит из нескольких блоков (таксономического, географического, библиографического, экологического). Для наполнения базы данных использованы литературные источники (в первую очередь, Ботанический журнал, Бюллетень МОИП, Новости сист. высш. растений и др.).
| Локальная флора | Пасвик1 | Ортина | Ковда | Пинега | Котлас | Вельск | Войсковицы | Мережа | ||||||||
| Регион | Мурманская обл. | Коми | Карелия | Архангельская обл. | Архангельская обл. | Архангельская обл. | Ленинградская обл. | Вологодская обл. | ||||||||
| Координаты | 69°13' с.ш. 29°24' в.д. | 67°45' с.ш. 54°8' в.д. | 66°46' с.ш. 32°45' в.д. | 64°40' с.ш. 43°20' в.д. | 61°16' с.ш. 46°23' в.д. | 61°10' с.ш. 42°7' в.д. | 59°45' с.ш. 29°50' в.д. | 59°31' с.ш. 36°32' в.д. | ||||||||
| Фракция | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S |
| Число видов | 368 | 285 | 237 | 220 | 430 | 374 | 628 | 614 | 589 | 564 | 571 | 556 | 637 | 607 | 567 | 553 |
| Число семейств | 62 | 55 | 51 | 50 | 72 | 70 | 82 | 81 | 87 | 87 | 90 | 89 | 91 | 88 | 95 | 93 |
| Asteraceae | 3 | 3 | 2 | 2 | 3 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Poaceae | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Cyperaceae | 2 | 1 | 3 | 3 | 2 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Caryophyllaceae | 4 | 4-5 | 4-5 | 5 | 5 | 5-6 | 4-6 | 6 | 6 | 5-7 | 7 | 7 | 8-9 | 5-7 | 4 | 4-5 |
| Brassicaceae | 5 | 13 | 9 | 9 | 4 | 5-7 | 8 | 8 | 5 | 8-9 | 9 | 9 | ||||
| Rosaceae | 7-8 | 9 | 6 | 6-8 | 4 | 4 | 4-6 | 4-5 | 5 | 4 | 4 | 4 | 4 | 4 | 5 | 4-5 |
| Scrophulariaceae | 9 | 6 | 10-11 | 10 | 6-7 | 5-6 | 8 | 7 | 7 | 5-7 | 5 | 5 | 8-9 | 5-7 | 6-7 | 7 |
| Fabaceae | 7-8 | 12 | 8-9 | 7 | 8 | 8 | 8-9 | 9 | 9-10 | 7 | 5-7 | 6-7 | 6 | |||
| Ranunculaceae | 10-11 | 10 | 7-8 | 6-8 | 6-7 | 7 | 4-6 | 4-5 | 9 | 8-9 | 6 | 6 | 10 | 10 | 8 | 8 |
| Lamiaceae | 10 | 10 | 6 | 8-9 | ||||||||||||
| Apiaceae | 10-12 | 9-10 | ||||||||||||||
| Salicaceae | 7-8 | 9 | 9 | 10 | 10 | 10 | 10 | 9-10 | ||||||||
| Polygonaceae | 10-11 | 11 | 10-11 | 10-12 | 10 | 10 | 10 | 10 | ||||||||
| Juncaceae | 6 | 4-5 | 4-5 | 4 | 8-9 | 8 | ||||||||||
| Ericaceae | 7-8 | 7-8 | 6-8 | 10-12 | 9-10 | |||||||||||
Примечание: 1 – для флоры «Пасвик» учтены виды заповедной территории и ближайших окрестностей.
| Локальная флора | Мцинское болото | Пестово | Ропажи | Уром | Себеж | Окский з-к | НД полесье1 | ЦЧЗ (З)2 | Хомутов. з-к3 | |||||||||
| Регион | Ленинградская обл. | Новгородская обл. | Латвия | Удмуртия | Псковская обл. | Рязанская обл. | Брянская обл. | Курская обл. | Украина | |||||||||
| Координаты | 58°55' с.ш. 30°17' в.д. | 58°37' с.ш. 35°13' в.д. | 57°00' с.ш. 24°40' в.д. | 56°50' с.ш. 53°00' в.д. | 56°15' с.ш. 28°28' в.д. | 55°45' с.ш. 40°50' в.д. | 52°30' с.ш. 34°00' в.д. | 51°11' с.ш. 36°24' в.д. | 47°22' с.ш. 38°17' в.д. | |||||||||
| Фракция | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | Sa | S | S | Sa | Sa | S |
| Число видов | 605 | 583 | 563 | 542 | 741 | 706 | 645 | 619 | 628 | 610 | 825 | 758 | 870 | 803 | 716 | 649 | 588 | 550 |
| Число семейств | 96 | 94 | 89 | 89 | 96 | 92 | 94 | 92 | 98 | 97 | 104 | 100 | 110 | 105 | 99 | 90 | 77 | 71 |
| Asteraceae | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Poaceae | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Cyperaceae | 3 | 3 | 3 | 3 | 3 | 3 | 4 | 4 | 4 | 4 | 3 | 3 | 3 | 3 | 5 | 3 | ||
| Caryophyllaceae | 5 | 5 | 7-8 | 6-7 | 8 | 7 | 7 | 6 | 5 | 5-6 | 7 | 5-6 | 6 | 4-6 | 8 | 8 | 7 | 7 |
| Brassicaceae | 6 | 9-10 | 7-8 | 8 | 6-7 | 9 | 6 | 8 | 10 | 10 | 6 | 8 | 9 | 10 | 7 | 7 | 4 | 4 |
| Rosaceae | 4 | 4 | 4 | 4 | 4-5 | 5 | 3 | 3 | 6 | 5-6 | 4 | 4 | 5 | 4-6 | 4 | 4 | 3 | 3 |
| Scrophulariaceae | 7-8 | 6 | 5-6 | 5 | 9 | 8 | 8 | 7 | 7-9 | 7 | 8-9 | 7 | 8 | 7-8 | 9-10 | 9 | 8 | 8 |
| Fabaceae | 7-8 | 7 | 5-6 | 6-7 | 4-5 | 4 | 5 | 5 | 3 | 3 | 5 | 5-6 | 4 | 4-6 | 3 | 5-6 | 6 | 5 |
| Ranunculaceae | 9 | 8 | 9 | 9 | 6-7 | 6 | 9-10 | 9-10 | 7-9 | 8-9 | 10-11 | 9-10 | 9-10 | |||||
| Lamiaceae | 10 | 9-10 | 10 | 10 | 10 | 10 | 9-10 | 9-10 | 7-9 | 8-9 | 8-9 | 9 | 7 | 7-8 | 6 | 5-6 | 5 | 6 |
| Apiaceae | 10 | 10-11 | 10 | 9 | 9-10 | 10 | 9-10 | 9-10 | ||||||||||
Примечание: 1 – Неруссо-Деснянское Полесье, 2 – Зоринский участок Центрально-Черноземного заповедника, 3 – заповедник Хомутовская степь.
Среди изученных локальных флор пять являются флорами заповедных территорий (4 заповедника и 1 заказник), однако включение заповедных флор в анализ не повлияло на тенденции изменения числа адвентивных видов, а по числу заносных видов эти флоры не являются самыми бедными. Два заповедника организованы сравнительно недавно: Пасвик – в 1992 г., Зоринский участок Центрально-Черноземного заповедника – в 1998 г., поэтому значительное число адвентивных видов еще сохранились на территории этих заповедников. Кроме того, для Пасвика помимо флоры заповедной территории учтены виды ближайших окрестностей. Заповедник «Хомутовская степь» расположен на юго-востоке Украины, в регионе сильно трансформированном человеком и с высокой степенью фрагментации растительного покрова. Присутствие адвентивных видов для конкретных флор определялось только при наличии списка видов или известных по литературе соответствующих исследований для данной точки [2, 5, 7, 11, 14, 15, 16, 23, 24, 25, 26, 29]. Для Зоринского участка Центрально-Черноземного заповедника (ЦЧЗ) из списка адвентивных видов исключены интродуцированные виды, природные популяции которых встречаются в какой-либо другой части территории заповедника (Festuca pratensis, Trifolium pratense и некоторые другие виды).
Взаимосвязь между числом адвентивных и аборигенных видов, а также между адвентивными видами и широтой оценена с помощью коэффициента корреляции Кендалла.
Изменение числа адвентивных видов в локальных флорах вдоль широтного градиента. Для Восточной Европы изменение доли адвентивных видов с широтой демонстрирует следующие закономерности (рис. 1):
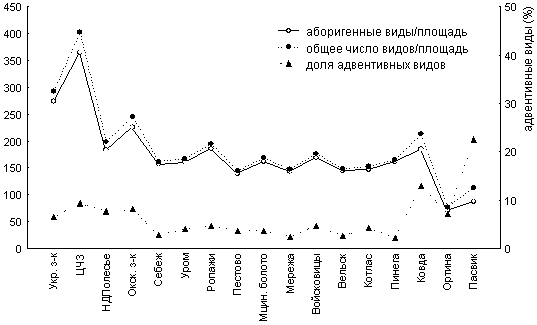
1. Если исключить тундровую зону, то, как видно из рисунка, число заносных видов в целом увеличивается с севера на юг, достигая максимума в зонах широколиственных лесов и лесостепи, затем немного уменьшается в степной зоне. Однако значимая корреляция между широтой и числом адвентивных видов (r=-0.23, p<0.18), а также их долей во флорах (r=0.05, p<0.76) не обнаружена. Тогда как видовое разнообразие флор зависит от широты (r=0.55, p<0.01) или точнее от факторов, меняющихся в широтном направлении. Отсутствие зависимости между адвентивными видами и широтой может объясняться как небольшой величиной выборки, так и ведущей ролью стохастических процессов, а также региональными особенностями антропогенного воздействия и особенностью биологии адвентивных видов.
2. Доля адвентивных видов больше в конкретных флорах тундровой зоны по сравнению с точками в прилегающих районах бореальной зоны. Аналогичная тенденция продемонстрирована и в других исследованиях. Так, Б. И. Груздев и В. А. Мартыненко [9] отметили, что процент участия адвентивных видов в конкретных флорах тундровой части Коми колеблется от 0 до 17, тогда как в таежных флорах адвентивная фракция не превышает 7%. Можно предложить несколько гипотез, объясняющих этот факт. Первая из них основана на «ненасыщенности» флоры северных территорий в результате воздействия ледникового покрова и необходимости длительного периода восстановления флоры. Другая более вероятная точка зрения связана с особенностями организации тундровых сообществ, а именно: при нарастании пессимума условий в высоких широтах происходит общее уменьшение видового разнообразия и разнообразия условий, увеличиваются экологические ниши видов и ослабляется конкуренция [19]. Третье объяснение связано с тем, что исследованные северные точки с более высоким, чем в бореальных флорах процентом адвентивных видов, относятся к подзоне южных тундр, где велика вероятность заноса значительного числа бореальных видов. К сожалению, сведения о локальных флорах из других частей тундровой зоны с полными списками адвентивных видов в них в литературе отсутствуют.
3. Не происходит резкого увеличения доли адвентивных видов в южном направлении, несмотря на бoльшую освоенность южных районов Восточной Европы и более длительный антропогенный пресс. Объяснение этого факта может быть связано с тем, что нарастание общего числа видов наблюдается на первых этапах антропогенного воздействия. В дальнейшем преобладает тенденция уменьшения видового богатства, причем этот процесс затрагивает также и адвентивные виды. Для регионов, издавна испытывавших значительное антропогенное влияние, отмечена сбалансированность видового состава флоры. Подобное явление, например, характерно для синантропной флоры Украины [21].
По результатам корреляционного анализа число адвентивных видов в локальных флорах Восточной Европы не зависит как от общего видового богатства локальных флор (r=0.26, p<0.14), так и числа аборигенных видов в них (r=0.19, p<0.28). Отсутствует корреляция также между долей адвентивных видов и разнообразием аборигенной фракции локальных флор, представляющим отношение числа видов к площади (r=0.09, p<0.55). Сведения о зависимости числа адвентивных видов от общего видового богатства в литературе довольно противоречивы. Кейз [31] изучал распределение интродуцированных видов птиц и обнаружил, что число интродуцированных видов на локальном уровне отрицательно связано с аборигенной авифауной. Однако говорить о влиянии конкуренции или других биотических взаимоотношениях в данном случае достаточно трудно, поскольку было выявлено, что число экзотов зависит также от общего числа заносных видов во всем регионе, т. е. велико региональное влияние. В некоторых исследованиях описана положительная зависимость числа заносных видов от общего видового богатства. Положительная корреляция была продемонстрирована между общим числом видов в каждом сообществе и процентом антропохоров в пойменных местообитаниях на юге Франции и в штате Орегон [34]. В 97 сообществах площадью 0.1 га, относящихся к 11 различным типам (штат Юта), число заносных видов было связано с видовым богатством аборигенной фракции [40]. Ранее этими же авторами были проведены более подробные исследования в различных по площади и растительным типам сообществах. Для четырех типов прерий (Колорадо, Вайоминг, Южная Дакота) число заносных видов на площадках 1 м2 уменьшалось с ростом видового богатства, на площадках 1000 м2 зависело от общего содержания азота в почве и покрытия аборигенных видов, а на ландшафтном уровне определялось высоким видовым богатством аборигенной фракции. В пяти лесных и луговых сообществах в горах Колорадо на всех уровнях число адвентивных видов зависело как от общего числа аборигенных видов, так и от других факторов. Был сделан вывод, что внедрение бoльшего числа заносных видов в точках с высоким разнообразием может объясняться уровнем доступных ресурсов в сообществах [39].
Систематическая структура локальных флор и ее изменения с учетом адвентивного компонента. Систематическая структура локальных флор и участие в ней ведущих семейств представлены в табл. 1 и 2. Для таксономических спектров исследуемых точек характерен довольно постоянный набор ведущих семейств, вариации которого определяются как широтой местности, так и конкретным местоположением локальной флоры.
Наиболее северной по спектру является флора бассейна реки Ортины (Большеземельская тундра), в систематической структуре которой лидирует сем. Poaceae, затем идут Asteraceae и Cyperaceae, на четвертом месте находится сем. Juncaceae. В спектре ведущих семейств присутствуют также лютиковые, которые занимают относительно высокое место, вересковые и ивовые. В целом арктический характер флоры подтверждается также отсутствием в первой десятке сем. Fabaceae и Lamiaceae (табл. 1). Большинство исследованных флор имеет бореальные черты с преобладанием сем. Asteraceae, Poaceae и Cyperaceae, а также Rosaceae и Caryophyllaceae на 4 и 5 местах таксономического спектра. Несколько наиболее южных по местоположению флор относятся к Среднеевропейской флористической области (табл. 2).
При анализе всей флоры, включая адвентивные виды, три ведущих семейства Asteraceae, Poaceae и Cyperaceae практически не меняют своего положения в структуре флор за исключением трех точек. Происходит только смена мест у злаков и осоковых в двух локальных флорах, расположенных в полосе редкостойных лесов (Пасвик и Ковда), а также осоки понижают свой ранг с третьего на пятый во флоре Зоринского участка Центрально-Черноземного заповедника. Высокий процент осоковых в целом не свойственен лесостепным и степным флорам, а их значительное участие во флоре Зоринского участка ЦЧЗ объясняется наличием болот на территории последнего.
В половине точек (8 локальных флор) отмечено увеличение процентного содержания и, соответственно, повышение ранга видов семейства Brassicaceae. Причем, как правило, повышение происходит не на один ранг, а на 2-4: с 5-7 места на 4 (Котлас), с 9 на 6 или 5 (Войсковицы, Мцинское болото, Ропажи) (табл. 1, 2). Увеличение роли семейства Brassicaceae может указывать на тенденцию «ксерофитизации» флор, поскольку присутствие видов этого семейства связано с увеличением количества однолетников-эфемеров. Аналогичная тенденция отмечена во флоре Мещеры: в аборигенной фракции флоры крестоцветные занимают шестое место, а во всей флоре с учетом адвентивных видов – третье [4]. Вместе с тем изменение статуса крестоцветных происходит не на всем широтном градиенте. В наиболее южных из исследованных флор сем. Brassicaceae не меняет своего местоположения в таксономическом спектре.
Достаточно стабильно положение семейств Rosaceae и Caryophyllaceae. С учетом участия адвентивных видов смена ранга у розоцветных происходит лишь в двух случаях: повышается статус семейства в Пасвике (с 9 на 7-8) и понижается в Котласе (с 4 на 5). Гвоздичные понижают свой ранг на один балл в четырех локальных флорах, причем смена мест происходит не за счет видов самого семейства, а в результате повышения роли видов семейства Brassicaceae. Бобовые существенно меняют свое местоположение лишь в двух точках (Ковда и Зоринский участок Центрально-Черноземного заповедника).
В пяти локальных флорах отмечены наибольшие изменения, которые, вероятно, определяются не широтным положением флор и принадлежностью к какому-либо биому, а скорее зависят от региональных особенностей и степени антропогенного воздействия. Все изменения касаются семейств, расположенных ниже трех первых мест. Сразу на несколько рангов меняют свой статус ряд семейств в спектре флоры заповедника Пасвик и его окрестностей. Здесь повышают свое место семейство Rosaceae (с 9 на 7,8), Fabaceae (с 12 на 7,8) и Brassicaceae (с 13 на 5). Существенно снизижается роль Juncaceae, Salicaceae и Ericaceae, значительное содержание видов которых характерно для арктических флор. В целом во флоре усилились бореальные черты. В четырех точках (Войсковицы, Ропажи, Уром, Окский заповедник) отмечены сходные изменения в систематической структуре флор. В их таксономических спектрах снизилась роль Scrophulariaceae и Caryophyllaceae, повысилась – Brassicaceae, а в Войсковицах – также и Lamiaceae.
В нескольких локальных флорах, различающихся по своей зональной принадлежности, нет изменений в систематической структуре, несмотря на варьирование содержания в них антропохоров. Остается постоянной структура локальной флоры Большеземельской тундры (южные гипоарктические тундры, 7.2% заносных видов), Вельска (средняя тайга, 2.6% заносных видов), Мережи (южная тайга, 2.5% заносных видов), Неруссо-Деснянского Полесья (зона широколиственных лесов, 7.7% заносных видов).
В целом изменение систематических структур флор при участии адвентивной фракции незначительно, происходит неравномерно и не зависит от широтного положения флоры. Вполне возможно также, что на анализ могла оказать влияние неполнота выявления видового состава флор и, в первую очередь, адвентивных видов. Изменение положения семейства Brassicaceae в нескольких точках явно недостаточно для подтверждения тенденции ксерофитизации флоры в результате заноса чужеродных видов.
Структура адвентивной фракции флор. Сравнение спектров семейств адвентивных фракций исследуемых локальных флор выявило, что они сильно различаются как по рангам семейств, так и по их набору. Наибольшим представительством обладает семейство Asteraceae, виды которого присутствуют во всех рассмотренных локальных флорах. Виды четырех семейств (Poaceae, Brassicaceae, Fabaceae, Lamiaceae) отмечены более чем в 75% точек, однако число видов в них сильно варьирует в различных точках. Следует отметить, что виды вышеперечисленных семейств, за исключением губоцветных, наиболее часто встречаются в мировой адвентивной флоре [36, 41].
Некоторые авторы предполагают наличие сходства в структурах адвентивной и аборигенной фракций. В работе В. Н. Голубева и И. В. Голубевой [8] указывается на близкое соотношение эколого-биологических признаков адвентивной и аборигенной флор Крыма, а именно: жизненных форм видов, типов вегетации, ритмов цветения, структурных типов корневой системы и пространственного размещения побегов. Соответствие в положении некоторых семейств, а именно, 2-3 ведущих, в систематических структурах нескольких точек по литературным данным отмечено М. В. Березуцким [3], а также при изучении адвентивной флоры Московской области [12]. Иная точка зрения высказана при анализе адвентивной флоры Чувашии: структура адвентивной флоры совершенно не соответствует структуре зональной флоры Чувашии или соседних регионов [10].
Таксономический анализ заносных видов в различных по широтному положению и антропогенному влиянию локальных флорах Восточной Европы показал, что не подтверждается мнение даже о частичном соответствии структуры адвентивной фракции структуре аборигенной фракции флоры. В аборигенной фракции всех рассмотренных локальных флор, за исключением трех северных, два первых места занимают семейства Asteraceae и Poaceae. В адвентивной флоре сложноцветные выходят на первое место лишь в 6 из 16 рассмотренных локальных флор. В шести локальных флорах на первом месте в структуре адвентивной фракции находятся крестоцветные, в трех случаях – злаки и бурачниковые. В одном случае по числу видов преобладают маревые (локальная флора Себеж) и бобовые (Зоринский участок ЦЧЗ), в двух локальных флорах маревые находятся на втором месте. Злаки занимают второе место в систематическом спектре адвентивной фракции лишь в трех локальных флорах в отличие от 14 аборигенных флор.
| N локальных флор | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 |
| Число заносных видов | 83 | 17 | 56 | 14 | 25 | 15 | 30 | 14 | 22 | 21 | 35 | 26 | 18 | 67 | 67 | 67 | 38 |
| Asteraceae | 10 | 2 | 8 | 3 | 5 | 2 | 3 | 2 | 2 | 1 | 7 | 4 | 2 | 13 | 12 | 7 | 9 |
| Poaceae | 9 | 7 | 8 | 1 | 2 | 1 | 1 | 2 | 4 | 1 | 3 | 1 | 1 | 8 | 3 | 9 | |
| Brassicaceae | 12 | 4 | 1 | 6 | 1 | 11 | 5 | 3 | 5 | 4 | 1 | 8 | 3 | 4 | 3 | ||
| Fabaceae | 9 | 1 | 5 | 2 | 1 | 1 | 2 | 1 | 3 | 1 | 1 | 1 | 6 | 7 | 10 | ||
| Boraginaceae | 2 | 3 | 4 | 6 | 5 | 2 | 1 | 2 | 1 | 1 | 1 | 2 | |||||
| Rosaceae | 5 | 1 | 6 | 2 | 3 | 2 | 5 | 7 | 3 | ||||||||
| Chenopodiaceae | 1 | 1 | 1 | 2 | 3 | 3 | 5 | 7 | 1 | 2 | |||||||
| Lamiaceae | 4 | 2 | 1 | 1 | 1 | 4 | 1 | 1 | 1 | 1 | 2 | 3 | 2 | ||||
| Apiaceae | 3 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | |||||||||
| Caryophyllaceae | 7 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | |||||
| Polygonaceae | 4 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | |||||||||
| Ranunculaceae | 3 | 2 | 1 | 1 | 1 |
Локальные флоры: 1 – Пасвик, 2 – Ортина, 3 – Ковда, 4 – Пинега, 5 – Котлас, 6 – Вельск, 7 – Войсковицы, 8 – Мережа, 9 – Мцинское болото, 10 – Пестово, 11 – Ропажи, 12 – Уром, 13 – Себеж, 14 – Окский з-к, 15 – Неруссо-Деснянское Полесье, 16 – Зоринский участок Центрально-Черноземного з-ка, 17 – Хомутовская степь з-к.
В большинстве точек таксономическая структура адвентивной фракции имеет более «южный» характер, что выражается или в преобладании, или в значительном участии в ней видов сем. Brassicaceae, Fabaceae и Boraginaceae (табл. 3). Подобная тенденция обнаружена при анализе адвентивной флоры Московской области [12].
На континентальном уровне явный широтный градиент в распределении числа адвентивных видов отсутствует. Доля адвентивных видов в ландшафте, если рассматривать конкретные флоры как флоры дискретных ландшафтов, не зависит от общего видового богатства флоры.
Добавление новых, заносных, видов не приводит к изменению основных закономерностей в распределении флор и не ведет к их повсеместной унификации. Интересно, что аналогичный вывод был сделан и при изучении городских флор, которые являются сильно трансформированными в антропогенном отношении и по числу адвентивных видов явно превосходят нативные флоры. Даже в пределах одного выдела (г. Москва), где, казалось бы, унификации флор ничто не препятствует, три участка разных железных дорог сильно различались и по флористическому составу, и по систематической структуре [4]. Причинами этого являются неравномерность антропогенного воздействия на флору, разные агенты и источники заноса, различия в режимах заноса видов [4], автохтонные особенности флоры, связанные с историей ее формирования, а также свойства самих видов. Следует учитывать также, что адвентивные виды обычно не играют заметной ценотической роли, встречаются единичными экземплярами, а наибольшее разнообразие заносных видов характерно для сообществ, расположенных вдоль железных и шоссейных дорог [18].
Не подтверждается мнение о соответствии систематической структуры адвентивной фракции флоры структуре аборигенной фракции. Таксономический состав адвентивной флоры сильно варьирует, но в целом имеет более южный характер по сравнению с аборигенной флорой. Вполне вероятно, что на результаты анализа могла оказать влияние неполнота выявления видового состава флор, тем более, что появление новых видов в каком-либо регионе и их дальнейшее присутствие во флоре – очень динамичный процесс.
Автор благодарит В. А. Бубыреву за материалы по локальным флорам Северо-запада России и Н. Г. Царевскую за участие в наполнении базы данных.
| |
кабинетом «Биоинформатики и моделирования биологических процессов» ИПЭЭ РАН. | |
| © 2004-. Кабинет "Биоинформатики и моделирования биологических процессов" ИПЭЭ РАН | |